|
|
| Line 66: |
Line 66: |
| | | | |
| | ==Structured Information== | | ==Structured Information== |
| − | {{JaponicaGene|
| |
| − | GeneName = Os07g0211500|
| |
| − | Description = Pericarp Color;red grain color gene|
| |
| − | Version = DQ204735.1 GI:78057266|
| |
| − | Length = 833bp|
| |
| − | Definition = Oryza sativa (japonica cultivar-group) cultivar H75 brown pericarp
| |
| − | and seed coat (Rc) gene, complete cds.|
| |
| − | Source = Oryza sativa Japonica Group (Japanese rice)
| |
| | | | |
| − | ORGANISM Oryza sativa Japonica Group
| |
| − | Eukaryota; Viridiplantae; Streptophyta; Embryophyta; Tracheophyta;
| |
| − | Spermatophyta; Magnoliophyta; Liliopsida; Poales; Poaceae; BEP
| |
| − | clade; Ehrhartoideae; Oryzeae; Oryza.|
| |
| − | Chromosome = [[:category:Japonica Chromosome 7|Chromosome 7]]|
| |
| − | AP = Chromosome 7|
| |
| − | source=RiceChromosome07
| |
| − | preset=GeneLocation
| |
| − | </gbrowseImage1>|
| |
| − | GSID = <gbrowseImage2>
| |
| − | name=NC_008397:19856181..19857859
| |
| − | source=RiceChromosome07
| |
| − | preset=GeneLocation
| |
| − | </gbrowseImage2>|
| |
| − | CDNA = <cdnaseq> 1 atggccggcg gcgaggcgca tgcggcgctg caggcggtgg cgcagagcct ccggtggacc
| |
| − | 61 tacagcctcc tctggcagct ctgcccccac caagggagctcg
| |
| − | 421 ctggtgtggg gggaggggca ctacaacggc gccgtcaaga cgcggaagtc gacggtgatg
| |
| − | 481 cagccgccgc cggcggagga ggaggacgac gccgaccacg cggcgcgcca ccggagccgg
| |
| − | 541 cagctgaggg agctctacga ctggctgcag caggccgggg agaactccag cggcggcgtg
| |
| − | 601 cagacgtcgt cgacgacggc gagccggcgg ccgggggcgg ctctgtcgcc ggaggacctg
| |
| − | 661 acggagacgg agtggttctt cctcatgtcg gcatcctact ccttccctcc cggcatcgggtta
| |
| − | 3421 cctggaaggg catttgcaag gagaggccat gtatggctca ctggagcaaa tgaagttgac
| |
| − | 3481 agcaaagtat tcctaagagc aattcttgcc aagacagttgtg tgcattcctg ttgtcgatgg cgtcctggaa attggaacta cggaaaaggtggaggaag atatgggcct gattcagtat gcaaggggca
| |
| − | 4201 tcttcatgga tcaacatggc atccacatga agcctaccct ctcacagcac tcaacatcca
| |
| − | 4261 acccagtcac ccactgtact catcagcatc caatccaggt tcagatgcaa ctaggtatca
| |
| − | 4321 ccagccaaac aaagtttgat tattcagatg agctcaatgc agatgaggag aatgatgaca
| |
| − | 4381 cagaagaaga gggcatgtca ggttcagaca ctaacaacac tgacactgaa aggaattcag
| |
| − | 4441 gccagctgca acttcaaatg caagaccaac tgaacatggt gagcaatgac caccagacaa
| |
| − | 4501 taccaaataa tgcagtttcc agtgagctaa tgcagtgtga gatgtcagaa gtggtaagag
| |
| − | 4561 atggctgctc aaataatatt ttagaggatg aaatccaaat gctgatggat tgccaaaaca
| |
| − | 4621 gtaattgtca gttaaatttg caagggccag atgagccttg tcactcttgg cattttctct
| |
| − | 4681 gcgaggagtt acaaaatgat taccagccagctactgaaga tcaagtggca tcacctgaaa atacccatta
| |
| − | 4921 cccaaaaaca ctcatgacaa tcctacatta caacacgctg cgacagcaag agatgaacat
| |
| − | 4981 caagaactac ttgccagttt cagagaaatc atcattctcc agatggacta ctcctgaagg
| |
| − | 5041 aagtgatgac aacaagacca tgatcagtcc aggcaccaca cagagaatgc tcaagagcat
| |
| − | 5101 cctgatgatt gttcccagta gtcactgcag ttacagggga gcagaaacac ctgaatcaag
| |
| − | 5161 gggcgggaaa ggcgcaagtg gaacgcgaaa agtcggtgcc atccaaggtg atttcagtgc
| |
| − | 5221 caaccatgtg ctgaaagaga ggagaagaag agagaagctc aatgagaagt tcataattct
| |
| − | 5281 gcgatctttg gtacctttca tgacaaagatggac aaggcgtcga tactaggcga
| |
| − | 6001 cacgatcgag tacgtgaagc agctaaggaa ccgcatacaa gagctcgagt cgtcgtcgtc
| |
| − | 6061 gtcgtcacga gcagccgccc gggcgccatc ggcggcggcc gccgggaggc ggaggaagag
| |
| − | 6121 atccgccgcc gccgccactg ccacggcggc ggaagggatg agcagcagca atggccgcaa
| |
| − | 6181 tggcggcgag gcggcggagg tggtgcaggt gtccatcatc gagagcgacg cgctgctgga
| |
| − | 6241 gctccggtgc ggttgcggcg gcggcggcgg cggtgtggtg ctgctccggg tgatgcaggc
| |
| − | 6301 gatgcaggag ctccagctgg aggtcaccgc cgtccaggcc tcgtgcgccg gtggcgagct
| |
| − | 6361 gctcgccgag ctgcgcgcca aggtcgtcgt tatgatcctg atctgcatga aaatgcaaat
| |
| − | 6421 gcaaatgcaa atgcagaatt aa</cdnaseq>|
| |
| − | AA = <aaseq>MAGGEAHAALQAVAQSLRWTYSLLWQLCPHQGSSLVWGEGHYNG
| |
| − | AVKTRKSTVMQPPPAEEEDDADHAARHRSRQLRELYDWLQQAGENSSGGVQTSSTTAS
| |
| − | RRPGAALSPEDLTETEWFFLMSASYSFPPGIGLPGRAFARRGHVWLTGANEVDSKVFL
| |
| − | RAILAKTVVCIPVVDGVLEIGTTEKVEEDMGLIQYARGIFMDQHGIHMKPTLSQHSTS
| |
| − | NPVTHCTHQHPIQVQMQLGITSQTKFDYSDELNADEENDDTEEEGMSGSDTNNTDTER
| |
| − | NSGQLQLQMQDQLNMVSNDHQTIPNNAVSSELMQCEMSEVVRDGCSNNILEDEIQMLM
| |
| − | DCQNSNCQLNLQGPDEPCHSWHFLCEELQNDYQPATEDQVASPENTHYPKTLMTILHY
| |
| − | NTLRQQEMNIKNYLPVSEKSSFSRWTTPEGSDDNKTMISPGTTQRMLKSILMIVPSSH
| |
| − | CSYRGAETPESRGGKGASGTRKVGAIQGDFSANHVLKERRRREKLNEKFIILRSLVPF
| |
| − | MTKMDKASILGDTIEYVKQLRNRIQELESSSSSSRAAARAPSAAAAGRRRKRSAAAAT
| |
| − | ATAAEGMSSSNGRNGGEAAEVVQVSIIESDALLELRCGCGGGGGGVVLLRVMQAMQEL
| |
| − | QLEVTAVQASCAGGELLAELRAKVVVMILICMKMQMQMQMQN"</aaseq>|
| |
| − | DNA = <dnaseqindica>1321..1386#668..1152#450..531# 1 atggccggcg gcgaggcgca tgcggcgctg caggcggtgg cgcagagcct ccggtggacc
| |
| − | 61 tacagcctcc tctggcagct ctgcccccac caagggtacc taccctacct acctacgaca
| |
| − | 121 cgatgcacag tgttcatcca tggccggcca tggcggatcg tcgtcgttgt cgatgatcat
| |
| − | 181 cgaaggaagc tagaggatat ggctcaatac tttgataata tatatactga tctctccgta
| |
| − | 241 caacaaaaat ataaaaattc tagctagtat cgaatgagac atatgctatg ctagtactac
| |
| − | 301 gaatctaaaa agatgtacat attttgattc gtattattag gatatatcac gagtttttat
| |
| − | 361 attttgagac ggatgtaata attctgaatt tagttgtgat ccatggcatg caggagctcg
| |
| − | 421 ctggtgtggg gggaggggca ctacaacggc gccgtcaaga cgcggaagtc gacggtgatg
| |
| − | 481 cagccgccgc cggcggagga ggaggacgac gccgaccacg cggcgcgcca ccggagccgg
| |
| − | 541 cagctgaggg agctctacga ctggctgcag caggccgggg agaactccag cggcggcgtg
| |
| − | 601 cagacgtcgt cgacgacggc gagccggcgg ccgggggcgg ctctgtcgcc ggaggacctg
| |
| − | 661 acggagacgg agtggttctt cctcatgtcg gcatcctact ccttccctcc cggcatcggg
| |
| − | 721 tatataataa aaaatataga tataaatatt taagcatgca tgcataaatt aaaccacact
| |
| − | 781 tcttgttacg tgttcttggc aaaatgatga acaattacca ctaattaatt ggagccagaa
| |
| − | 841 accctaaaga tttacccacc tggttaatta atcggtgtgt tgatccacgc atgcatgcat
| |
| − | 901 gcagaaaatc aagatcagga tagctccttt tcttttgcag gttaattagc tagatcttca
| |
| − | 961 cgtataatta gctagctaga ttttaaaata taatttattc aatttgattt atgattttta
| |
| − | 1021 ttttttattt caaatagata caactgtata caaaatttta ttttggtaca tacctccgat
| |
| − | 1081 ccaactacat cagaggtaaa aaaaaaatta aaccgttgga attgattaga acaagatcgt
| |
| − | 1141 gcggtcaaat tatatcataa ctaacttttc tgattctcta aagcatagag atgtatatat
| |
| − | 1201 acatcgtatt attaggctct atatttcctg attaacacta gatgcatata taattttgat
| |
| − | 1261 agtcaaaata tacttttgat aggctctaaa gaaaaactta ataacatgta ctccctccat
| |
| − | 1321 atacttttga tagtcatatt tcatcttgac acacagatca agtataagta attctactta
| |
| − | 1381 tcatccattt aaacacgcta ctagttattc ctcataaaca agcgattcat taatatttac
| |
| − | 1441 atttctcgat gcttgtgtag ccaatattgt gtggaagaat ggaatgtcat taagaggata
| |
| − | 1501 ggttgttgga ttgaaatatg cctatcaaaa ataaattttt agatttgaaa atatgcctat
| |
| − | 1561 caaaagtaga tggagggagt attaattaat gtgaatttcc aatcctactg ttgtgatatt
| |
| − | 1621 aggctttgta ccttcttgtc caggaggtat atatatggct cttttaagga tgggagaaaa
| |
| − | 1681 tatcatcttt aatacaacta tatatggctt ttgtttgata aatacaactt ttattttgta
| |
| − | 1741 tgaatacaaa tatattgata aatatccacc attataatcc taacccatta ggatcatatg
| |
| − | 1801 gtgtatattt ttttaactat ttgtttttta taaattaata ttaagagatc acaataaaaa
| |
| − | 1861 tatagtatta tgaaagtact cttaacaaca tatccaatga taaaattatt attattacaa
| |
| − | 1921 aatatagtgg tcaaattgta tagaattcaa tagcctgatt ttatgacgtc aagtaaatta
| |
| − | 1981 aataaagaat gaaggtagtg ctagagtgat caaacaatat ctctcctaaa atatgtccta
| |
| − | 2041 taagttttac tccataaatc caagggtcaa aagttgttgg gttatttttt tagataataa
| |
| − | 2101 catactaccc cttttcaaaa tgtatgattc tattgacttt ttgcacaaca tttaaccatt
| |
| − | 2161 tgtcatatta aaaattagta taaacatcta aaaatataag ttacaattat attttatttg
| |
| − | 2221 atgataaaac aactcacaac aaaataaata atatttatat aatctttttg gaataaaacg
| |
| − | 2281 aatgatcaaa cattattcaa aaagtcaatg gtatagtacg ttttgaaatt gatagactat
| |
| − | 2341 gagagcaaaa ttttgagata acatggaaaa ttatcctctt agacattgca ctgtgtaata
| |
| − | 2401 attaataata atgaatgaaa ggctaagact tttcttccac cttatataag tggttgaata
| |
| − | 2461 tatagcaatc acatcattac atgattttgt aaccaaccgt ctctatagct ccgatacagt
| |
| − | 2521 gctagtttca catcgtaata attaaagagt ataataataa atcgaggtgt acttctcatc
| |
| − | 2581 gatgaagtga tgtgccgctt agctaaatta aactcgtatg cgaaaaatca gtatatgtcc
| |
| − | 2641 ggttaatttc taagagagag attgagagag aataattgcg cccctccaaa tccccctctt
| |
| − | 2701 ggacgttagg gagctatata gacggtattg ctaagtgcga tgtgtacata acgtacctgt
| |
| − | 2761 cgtaggaaca tttctcatcc aaattaagta gtaatgcatg gcatgaaatc catttttgta
| |
| − | 2821 ttttgcatgg caaagaatga caacaaggaa tacactagct agccctgccc tttttcaatt
| |
| − | 2881 taatttaaca tcaaacttag ttattgtatt tcttttgtca gaatagcatg cattgcatac
| |
| − | 2941 tctttaaaaa taattaatta gtgtatttta ctagtcttac aaaagtatca agagagacaa
| |
| − | 3001 ctaattatag ttgggagaca ccaaacttgt ttttaataat gacaattaaa accctacctc
| |
| − | 3061 tacatccaac atagacgtac atagtccgaa ggcgccaaat atttgtacat ttagctacca
| |
| − | 3121 gatttcagta cgagttctca cattataatt ttgatttttt tatttttttt ataaacaatc
| |
| − | 3181 tggtaccctt ttatgtctgg aaggaaaaaa aaaatctaaa ttgcaacatt ttagtcggtg
| |
| − | 3241 agaatggtac tctgtcctag ctactttcta cacatgagag agagagagag agagagagag
| |
| − | 3301 agagccttta attgcccttg cccatgcatc tttctttgca cacatgtatg cttttcacat
| |
| − | 3361 tgtcatgagg agagaacttg ttaagttgca cacatgtgtg ctttgcatgt cttcaggtta
| |
| − | 3421 cctggaaggg catttgcaag gagaggccat gtatggctca ctggagcaaa tgaagttgac
| |
| − | 3481 agcaaagtat tcctaagagc aattcttgcc aaggttcagc catcaccttc tcttacctat
| |
| − | 3541 ttttcactct gaatgccaac agtgctttgc acattgtagt ctgtttgcag actgcaaatg
| |
| − | 3601 atgaccataa tcagatcaga aaataaaata atattatata ctttttgagc cagctagcaa
| |
| − | 3661 gaatatgtaa caataattct cctttttttt tcttgttctt ttccctgatg tggtgcataa
| |
| − | 3721 caaataacca aactgatgaa tggcagagtg ctggtatcca ggtatttgcc tctaaaagta
| |
| − | 3781 gctacacgtt tactatgaaa ttttgtggct tttgttcatc tttggatgca gtggccatta
| |
| − | 3841 tctaaaaact atgaatttcc agactgcagt ttttatctaa ttttgtgact ttgtacatca
| |
| − | 3901 gacagttgtg tgcattcctg ttgtcgatgg cgtcctggaa attggaacta cggaaaaggt
| |
| − | 3961 gatttcgtat attatcagct gacaatctaa ttatatgggc catataatta agtataaatc
| |
| − | 4021 aaaatacctc ataatatatt ataaagtatc taatgtgatt atgtgaatat tggctatttc
| |
| − | 4081 aatgtaattt gatatatgaa actgataatc ctctgaaact ccgtaaggat caaactaatc
| |
| − | 4141 aaaatgtata tattttcaag gtggaggaag atatgggcct gattcagtat gcaaggggca
| |
| − | 4201 tcttcatgga tcaacatggc atccacatga agcctaccct ctcacagcac tcaacatcca
| |
| − | 4261 acccagtcac ccactgtact catcagcatc caatccaggt tcagatgcaa ctaggtatca
| |
| − | 4321 ccagccaaac aaagtttgat tattcagatg agctcaatgc agatgaggag aatgatgaca
| |
| − | 4381 cagaagaaga gggcatgtca ggttcagaca ctaacaacac tgacactgaa aggaattcag
| |
| − | 4441 gccagctgca acttcaaatg caagaccaac tgaacatggt gagcaatgac caccagacaa
| |
| − | 4501 taccaaataa tgcagtttcc agtgagctaa tgcagtgtga gatgtcagaa gtggtaagag
| |
| − | 4561 atggctgctc aaataatatt ttagaggatg aaatccaaat gctgatggat tgccaaaaca
| |
| − | 4621 gtaattgtca gttaaatttg caagggccag atgagccttg tcactcttgg cattttctct
| |
| − | 4681 gcgaggagtt acaaaatgat taccagccag gtattacatt tgagaagata atccttcaaa
| |
| − | 4741 agcacccttg ttccaaaaat atatatttgt actcttcaca caagcactgc catttttttt
| |
| − | 4801 cttttttgca tacatcctca attcttgcat ttcttttcca tatatttgat acaactgtct
| |
| − | 4861 ccatttccct tctgtcacag ctactgaaga tcaagtggca tcacctgaaa atacccatta
| |
| − | 4921 cccaaaaaca ctcatgacaa tcctacatta caacacgctg cgacagcaag agatgaacat
| |
| − | 4981 caagaactac ttgccagttt cagagaaatc atcattctcc agatggacta ctcctgaagg
| |
| − | 5041 aagtgatgac aacaagacca tgatcagtcc aggcaccaca cagagaatgc tcaagagcat
| |
| − | 5101 cctgatgatt gttcccagta gtcactgcag ttacagggga gcagaaacac ctgaatcaag
| |
| − | 5161 gggcgggaaa ggcgcaagtg gaacgcgaaa agtcggtgcc atccaaggtg atttcagtgc
| |
| − | 5221 caaccatgtg ctgaaagaga ggagaagaag agagaagctc aatgagaagt tcataattct
| |
| − | 5281 gcgatctttg gtacctttca tgacaaaggt aattaagtac tccctctatt tctataaagc
| |
| − | 5341 cgtatttgac tagttatctt atttagaaag tatgtgcaaa tatgtaaaat ataagtcata
| |
| − | 5401 cttaaagaac ttttaatgtt attaaataat aagtcacacc aaaaataaaa catatatatt
| |
| − | 5461 tttaataaga caaatgatta aatgtatata taaaaattaa tagcgtcaca tattttaaaa
| |
| − | 5521 tagaggggta tttaagtacc cacaggatca tcaaaattca gttatctttt cttaagcctc
| |
| − | 5581 taacgaacat tggaagatcc tcactaatgg cagcatgaat ctagggttca ctatttcgga
| |
| − | 5641 atgcaaaata tgttttaccg ggcatccgat ttttaaaaaa ttcagaatga agaaaattga
| |
| − | 5701 atctttttta tggatttgaa taaatcttga taaattcgaa aaaatttccg aacttttggc
| |
| − | 5761 cagaagtgaa tcctacccgt atccaccggt aataaaccta aatttttggg agtaatgaat
| |
| − | 5821 taatgttata tataatccat gaattatata gttccaaact actccgtaac aaattttcag
| |
| − | 5881 gagtagtgaa attaatatta ttacaatctc agaaaaaaat ggcagaaaca attaatctgt
| |
| − | 5941 tttcaattat taattaattt gtttttgtgt ccagatggac aaggcgtcga tactaggcga
| |
| − | 6001 cacgatcgag tacgtgaagc agctaaggaa ccgcatacaa gagctcgagt cgtcgtcgtc
| |
| − | 6061 gtcgtcacga gcagccgccc gggcgccatc ggcggcggcc gccgggaggc ggaggaagag
| |
| − | 6121 atccgccgcc gccgccactg ccacggcggc ggaagggatg agcagcagca atggccgcaa
| |
| − | 6181 tggcggcgag gcggcggagg tggtgcaggt gtccatcatc gagagcgacg cgctgctgga
| |
| − | 6241 gctccggtgc ggttgcggcg gcggcggcgg cggtgtggtg ctgctccggg tgatgcaggc
| |
| − | 6301 gatgcaggag ctccagctgg aggtcaccgc cgtccaggcc tcgtgcgccg gtggcgagct
| |
| − | 6361 gctcgccgag ctgcgcgcca aggtcgtcgt tatgatcctg atctgcatga aaatgcaaat
| |
| − | 6421 gcaaatgcaa atgcagaatt aa
| |
| − | </dnaseqindica>|
| |
| − | Link = [http://www.ncbi.nlm.nih.gov/nuccore/78057269]|
| |
| − | }}
| |
| | [[Category:Genes]] | | [[Category:Genes]] |
| | [[Category:Oryza Sativa Japonica Group]] | | [[Category:Oryza Sativa Japonica Group]] |
Rc is a domestication-related gene required for red pericarp in rice (Oryza sativa) and it encodes a basic helix-loop-helix (bHLH) protein that was fine-mapped to an 18.5-kb region on rice chromosome 7 using a cross between Oryza rufipogon (red pericarp) and O. sativa cv Jefferson (white pericarp).
characteristic
Different colors, such as purple, brown, red and white, occur in the pericarp of rice. Here, two genes affecting proanthocyanidin synthesis in red- and brown-colored rice were elucidated, one of the gene is Rc.[1])
Mutation

picture1. Length mutation (from reference [2]).  Grain color has been the target of selection in domesticated Asian rice (as well as other domesticated grains), and the vast majority of O. sativa varieties have white (nonpigmented) grains, compared to the dark red grains that characterize the wild relative and other wild species. The genetic basis of white grains in O. sativa has been pinpointed to loss-of-function mutations in the Rc gene, a regulatory protein in the proanthocyanidin synthesis pathway; these loss-of-function mutations prevent the development of a pigmented pericarp layer. A survey of 337 white pericarp rice cultivars showed that the majority (>97%) have a 14-bp deletion in exon 7 of the gene, resulting in a premature stop codon and a non-functional allele referred to as rc.
Our findings for the pericarp color gene, Rc, indicate that remarkably similar genetic mechanisms can be at play. We find that two of the three white pericarp O. glaberrima varieties in our sample harbor a unique point mutation that is predicted to result in a premature stop codon in exon 7 of the Rc gene.
Instead, two samples contained a novel point mutation predicted to result in a premature stop-codon in exon 7, hereafter referred to as allele rc-g1 . This mutation is 146 bp upstream of the Rc-s point mutation and 201 bp upstream of the initiation of the 14-bp rc deletion. The third white pericarp O. glaberrima contained no identifiable insertions, deletions, or premature stop-codons, nor does it differ from the red-pericarp O. glaberrima sequences for any predicted amino-acid changes. Comparison of this sample to the most closely related red-pericarp sample showed that they were identical for the sequenced region upstream of the start codon (>1.5 kb) and for the first intron; both of these regions are potentially important as cis-regulatory regions in plants.
This study also shows that white pericarps might be achieved by mutations in cis-regulatory regions of Rc or potentially mutations in other genes, a pattern that was not observed in an extensive survey of O. sativa cultivars.(from reference [2])
Crop domestication can serve as a model of plant evolutionary processes. It involves a series of selection events from standing natural variation and newly occurring mutations and combinations of mutations as a result of natural crossings in populations during local adaptation and propagation of plant lines to other cultivation areas.[3])
To determine the subpopulation origin of the Rc mutations,we examined ancestral haplotypes across the Rc coding sequence and promoter region in 103 genetically diverse, redgrained rices. Four rice insertion/deletion polymorphism (RID) and two rice microsatellite (RM) polymorphisms were used to construct haplotypes across the 6.5-kb region containing the Rc gene.[4])
Grain color has been the target of selection in domesticated Asian rice (as well as other domesticated grains), and the vast majority of O. sativa varieties have white (nonpigmented) grains, compared to the dark red grains that characterize the wild relative and other wild species. The genetic basis of white grains in O. sativa has been pinpointed to loss-of-function mutations in the Rc gene, a regulatory protein in the proanthocyanidin synthesis pathway; these loss-of-function mutations prevent the development of a pigmented pericarp layer. A survey of 337 white pericarp rice cultivars showed that the majority (>97%) have a 14-bp deletion in exon 7 of the gene, resulting in a premature stop codon and a non-functional allele referred to as rc.
Our findings for the pericarp color gene, Rc, indicate that remarkably similar genetic mechanisms can be at play. We find that two of the three white pericarp O. glaberrima varieties in our sample harbor a unique point mutation that is predicted to result in a premature stop codon in exon 7 of the Rc gene.
Instead, two samples contained a novel point mutation predicted to result in a premature stop-codon in exon 7, hereafter referred to as allele rc-g1 . This mutation is 146 bp upstream of the Rc-s point mutation and 201 bp upstream of the initiation of the 14-bp rc deletion. The third white pericarp O. glaberrima contained no identifiable insertions, deletions, or premature stop-codons, nor does it differ from the red-pericarp O. glaberrima sequences for any predicted amino-acid changes. Comparison of this sample to the most closely related red-pericarp sample showed that they were identical for the sequenced region upstream of the start codon (>1.5 kb) and for the first intron; both of these regions are potentially important as cis-regulatory regions in plants.
This study also shows that white pericarps might be achieved by mutations in cis-regulatory regions of Rc or potentially mutations in other genes, a pattern that was not observed in an extensive survey of O. sativa cultivars.(from reference [2])
Crop domestication can serve as a model of plant evolutionary processes. It involves a series of selection events from standing natural variation and newly occurring mutations and combinations of mutations as a result of natural crossings in populations during local adaptation and propagation of plant lines to other cultivation areas.[3])
To determine the subpopulation origin of the Rc mutations,we examined ancestral haplotypes across the Rc coding sequence and promoter region in 103 genetically diverse, redgrained rices. Four rice insertion/deletion polymorphism (RID) and two rice microsatellite (RM) polymorphisms were used to construct haplotypes across the 6.5-kb region containing the Rc gene.[4])
Function
 Rc alleles from the temperate ecotype of weedy lines LD and C9541 could represent the functional alleles most distant to those in group I (Figure 4). All the donor lines of weedy and cultivated rice in both groups were identified as dormant genotypes, or dormancy was associated with the Rc locus in our previous or present research. Thus, phylogenetic and dormancy data suggest that Rc alleles functional for pericarp color most likely also function for seed dormancy.[5])
Rc alleles from the temperate ecotype of weedy lines LD and C9541 could represent the functional alleles most distant to those in group I (Figure 4). All the donor lines of weedy and cultivated rice in both groups were identified as dormant genotypes, or dormancy was associated with the Rc locus in our previous or present research. Thus, phylogenetic and dormancy data suggest that Rc alleles functional for pericarp color most likely also function for seed dormancy.[5])
Expression
RT-PCR experiments confirmed that the Rc gene was expressed in both red- and white-grained rice but that a shortened transcript was present in white varieties.We anticipated that similar expression levels of Rc would be detected in red and white seeds. Our results confirmed this expectation and further demonstrated that the RNA transcript from cv Jefferson contained the 14-bp deletion predicted from the sequence information.(from reference [6])
Nucleotide Polymorphisms
Phylogenetic analysis, supported by comparative mapping in rice and maize (Zea mays), showed that Rc, a positive regulator of proanthocyanidin, is orthologous with INTENSIFIER1, a negative regulator of anthocyanin production in maize, and is not in the same clade as rice bHLH anthocyanin regulators.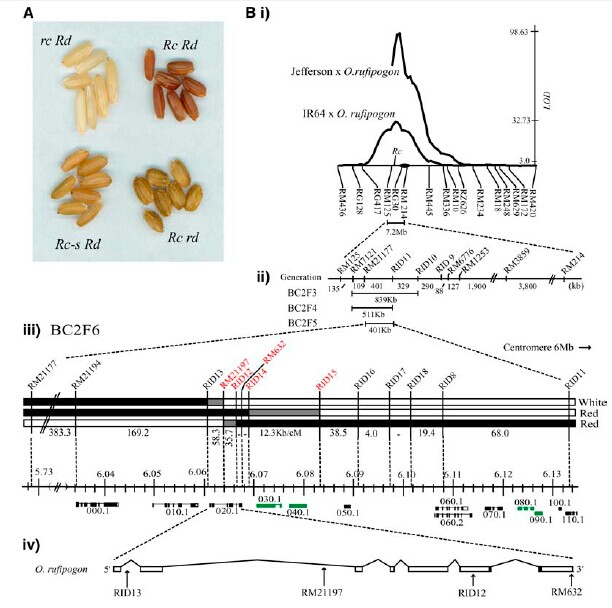 (from reference [6])
(from reference [6])
Allele Distribution
Sequencing of the alleles from both mapping parents as well as from two independent genetic stocks of Rc revealed that the dominant red allele differed from the recessive white allele by a 14-bp deletion within exon 6 that knocked out the bHLH domain of the protein.(from reference [6])
Recombination
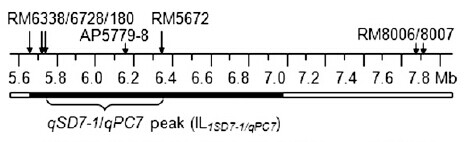
The rg7.1 locus was originally mapped to a 7.2-Mb region that included the centromere on rice chromosome 7. Given the low frequency of recombination across this region, it was not clear whether positional cloning would be feasible. This study demonstrates that even in regions that are recombinationally repressed, map-based gene isolation offers a viable approach.(from reference [6])
Evolution
Repeated phenotypic evolution can occur at the interspecific level, where it is manifest as the appearance of the same trait in multiple domesticated species, and at the intraspecific level, where the same trait arises multiple times within a single crop.
In rice, the opportunity exists to examine the genetic basis of repeated trait evolution at both the interspecific and intraspecific level.
At the intraspecific level, repeated phenotypic evolution during domestication can be explored in two contexts; one is the repeated evolution of a trait in varieties resulting from a single domestication event, the other is repeated evolution of a trait in varieties that result from multiple, independent domestication events within the same species.(from reference [2])
Labs working on this gene
- Department of Biology, Washington University in St. Louis, St. Louis, Missouri, USA
- Plant Science Department, South Dakota State University, Brookings, South Dakota 57007, †Biosciences Research Laboratory, U.S.
- Department of Agriculture–Agricultural Research Service, Fargo, North Dakota 58105, ‡Northern Crop Science Laboratory, U.S.
- Department of Agriculture–Agricultural Research Service, Fargo, North Dakota 58102, §National Institute of Agrobiologica
- Sciences, Tsukuba, Ibaraki 305-8602, Japan
- Agricultural College, Yangzhou University, Yangzhou 225008, China
- Department of Plant Breeding and Genetics, Cornell University, Ithaca, New York 14953-1901
- Department of Plant Biology, Cornell University, Ithaca, New York 14853
- Department of Plant Breeding and Genetics, Cornell University, Ithaca, New York, United States of America
- International Rice Research Institute, Los Ban˜ os, Philippines*
- Indonesian Center for Agricultural Biotechnology and Genetic Resources Research and Development, Bogor, Indonesia
- Department of Agronomy, Chungbuk National University, Chongju, Republic of Korea
- National Institute of Agricultural Biotechnology, Suwon, Republic of Korea
- Department of Biological Statistics and Computational Biology, Cornell University, Ithaca, New York, United States of America
- Plant Genome Research Unit, National Institute of Agrobiological Sciences, 2-1-2 Kannondai, Tsukuba, Ibaraki, 305-8602 Japan
- QTL Genomics Research Center, National Institute of Agrobiological Sciences, 2-1-2 Kannondai, Tsukuba, Ibaraki, 305-8602 Japan
- Genetic Diversity Department, National Institute of Agrobiological Sciences, Tsukuba, Ibaraki 305-8602, Japan
- Department of Biological Science and Technology, Tokyo University of Science, Noda, Chiba 278-8510, Japan
- Research Institute for Biresources, Okayama University, Kurashiki, Okayama 710-0046, Japan
- National Agricultural Research Center for Kyushu Okinawa Region, National Agriculture and Bio-oriented Research Organization, Nishigoshi, Kikuchi, Kumamoto 861-1192, Japan
- National Institute for Basic Biology, Okazaki, Aichi 444-8585, Japan
- Plant Breeding Laboratory, Graduate School of Agriculture, Hokkaido University, Sapporo, Hokkaido 060-8589, Japan
References
- ↑ Tsutomu Furukawa, Masahiko Maekawa, Tomoyuki Oki, Ikuo Suda, Shigeru Iida, Hiroaki Shimada, Itsuro Takamure, Koh-ichi Kadowaki The Rc and Rd genes are involved in proanthocyanidin synthesis in rice pericarp The Plant Journal, 2006, 49(1): 91-102.
- ↑ 2.0 2.1 2.2 Xing-You Gu; Michael E. Foley; David P. Horvath; James V. Anderson; Jiuhuan Feng; Lihua Zhang; Chase R. Mowry; Heng Ye; Jeffery C. Suttle; Koh-ichi Kadowaki; Zhongxiang Chen. Association Between Seed Dormancy and Pericarp Color Is Controlled by a Pleiotropic Gene That Regulates Abscisic Acid and Flavonoid Synthesis in Weedy Red Rice;Genetics, 2011, 189(4): 1515-1524.
- ↑ Saeko Konishi, Kaworu Ebana and Takeshi Izawa Inference of the japonica Rice Domestication Process from the Distribution of Six Functional Nucleotide Polymorphisms of Domestication-Related Genes in Various Landraces and Modern Cultivars Plant and Cell Physiology, 2008, 49(9): 1283-1293.
- ↑ Megan T. Sweeney; Michael J. Thomson; Yong Gu Cho; Yong Jin Park; Scott H. Williamson; Carlos D. Bustamante; Susan R. McCouch Global Dissemination of a Single Mutation Conferring White Pericarp in Rice PLoS Genetics, 2007, 3(8): 1418-1424.
- ↑ B. L. GROSS, F. T. STEFFEN, K. M. OLSEN The molecular basis of white pericarps in African domesticated rice: novel mutations at the Rc gene.Journal of Evolutionary Biology, 2010, 23(12): 2747-2753.
- ↑ 6.0 6.1 6.2 6.3 Megan T. Sweeney, Michael J. Thomson, Bernard E. Pfeil, Susan McCouch Caught Red-Handed: Rc Encodes a Basic Helix-Loop-Helix Protein Conditioning Red Pericarp in Rice The Plant Cell, 2006, 18(2): 283-294.
Structured Information

 Grain color has been the target of selection in domesticated Asian rice (as well as other domesticated grains), and the vast majority of O. sativa varieties have white (nonpigmented) grains, compared to the dark red grains that characterize the wild relative and other wild species. The genetic basis of white grains in O. sativa has been pinpointed to loss-of-function mutations in the Rc gene, a regulatory protein in the proanthocyanidin synthesis pathway; these loss-of-function mutations prevent the development of a pigmented pericarp layer. A survey of 337 white pericarp rice cultivars showed that the majority (>97%) have a 14-bp deletion in exon 7 of the gene, resulting in a premature stop codon and a non-functional allele referred to as rc.
Our findings for the pericarp color gene, Rc, indicate that remarkably similar genetic mechanisms can be at play. We find that two of the three white pericarp O. glaberrima varieties in our sample harbor a unique point mutation that is predicted to result in a premature stop codon in exon 7 of the Rc gene.
Instead, two samples contained a novel point mutation predicted to result in a premature stop-codon in exon 7, hereafter referred to as allele rc-g1 . This mutation is 146 bp upstream of the Rc-s point mutation and 201 bp upstream of the initiation of the 14-bp rc deletion. The third white pericarp O. glaberrima contained no identifiable insertions, deletions, or premature stop-codons, nor does it differ from the red-pericarp O. glaberrima sequences for any predicted amino-acid changes. Comparison of this sample to the most closely related red-pericarp sample showed that they were identical for the sequenced region upstream of the start codon (>1.5 kb) and for the first intron; both of these regions are potentially important as cis-regulatory regions in plants.
This study also shows that white pericarps might be achieved by mutations in cis-regulatory regions of Rc or potentially mutations in other genes, a pattern that was not observed in an extensive survey of O. sativa cultivars.(from reference [2])
Crop domestication can serve as a model of plant evolutionary processes. It involves a series of selection events from standing natural variation and newly occurring mutations and combinations of mutations as a result of natural crossings in populations during local adaptation and propagation of plant lines to other cultivation areas.[3])
To determine the subpopulation origin of the Rc mutations,we examined ancestral haplotypes across the Rc coding sequence and promoter region in 103 genetically diverse, redgrained rices. Four rice insertion/deletion polymorphism (RID) and two rice microsatellite (RM) polymorphisms were used to construct haplotypes across the 6.5-kb region containing the Rc gene.[4])
Grain color has been the target of selection in domesticated Asian rice (as well as other domesticated grains), and the vast majority of O. sativa varieties have white (nonpigmented) grains, compared to the dark red grains that characterize the wild relative and other wild species. The genetic basis of white grains in O. sativa has been pinpointed to loss-of-function mutations in the Rc gene, a regulatory protein in the proanthocyanidin synthesis pathway; these loss-of-function mutations prevent the development of a pigmented pericarp layer. A survey of 337 white pericarp rice cultivars showed that the majority (>97%) have a 14-bp deletion in exon 7 of the gene, resulting in a premature stop codon and a non-functional allele referred to as rc.
Our findings for the pericarp color gene, Rc, indicate that remarkably similar genetic mechanisms can be at play. We find that two of the three white pericarp O. glaberrima varieties in our sample harbor a unique point mutation that is predicted to result in a premature stop codon in exon 7 of the Rc gene.
Instead, two samples contained a novel point mutation predicted to result in a premature stop-codon in exon 7, hereafter referred to as allele rc-g1 . This mutation is 146 bp upstream of the Rc-s point mutation and 201 bp upstream of the initiation of the 14-bp rc deletion. The third white pericarp O. glaberrima contained no identifiable insertions, deletions, or premature stop-codons, nor does it differ from the red-pericarp O. glaberrima sequences for any predicted amino-acid changes. Comparison of this sample to the most closely related red-pericarp sample showed that they were identical for the sequenced region upstream of the start codon (>1.5 kb) and for the first intron; both of these regions are potentially important as cis-regulatory regions in plants.
This study also shows that white pericarps might be achieved by mutations in cis-regulatory regions of Rc or potentially mutations in other genes, a pattern that was not observed in an extensive survey of O. sativa cultivars.(from reference [2])
Crop domestication can serve as a model of plant evolutionary processes. It involves a series of selection events from standing natural variation and newly occurring mutations and combinations of mutations as a result of natural crossings in populations during local adaptation and propagation of plant lines to other cultivation areas.[3])
To determine the subpopulation origin of the Rc mutations,we examined ancestral haplotypes across the Rc coding sequence and promoter region in 103 genetically diverse, redgrained rices. Four rice insertion/deletion polymorphism (RID) and two rice microsatellite (RM) polymorphisms were used to construct haplotypes across the 6.5-kb region containing the Rc gene.[4])
 Rc alleles from the temperate ecotype of weedy lines LD and C9541 could represent the functional alleles most distant to those in group I (Figure 4). All the donor lines of weedy and cultivated rice in both groups were identified as dormant genotypes, or dormancy was associated with the Rc locus in our previous or present research. Thus, phylogenetic and dormancy data suggest that Rc alleles functional for pericarp color most likely also function for seed dormancy.[5])
Rc alleles from the temperate ecotype of weedy lines LD and C9541 could represent the functional alleles most distant to those in group I (Figure 4). All the donor lines of weedy and cultivated rice in both groups were identified as dormant genotypes, or dormancy was associated with the Rc locus in our previous or present research. Thus, phylogenetic and dormancy data suggest that Rc alleles functional for pericarp color most likely also function for seed dormancy.[5])
 (from reference [6])
(from reference [6])